
生长素是最重要的植物激素,主要合成部位是芽、幼嫩的叶和发育中的种子,通过极性运输分配到植物各处组织,调控生长发育——低浓度生长素促进生长,高浓度生长素抑制生长。生长素的极性运输对生长素的分配至关重要。那么,生长素极性运输是如何实现的呢?
近日,浙江大学医学院生物物理系长聘副教授/附属第四医院双聘教授郭江涛指导的浙江大学-湖北大学联合研究团队阐明了生长素转运蛋白PIN介导生长素极性运输的分子机制。这项工作于北京时间8月2日刊登在国际顶级期刊《自然》上。该成果作为植物生长素极性运输研究的重大突破,解决了植物向性这一个百年科学难题中的关键一环,为人们进一步调控生长素极性运输奠定了基础。
生长素及其转运蛋白PIN的发现过程
19世纪末,英国著名生物学家、进化论的奠基人达尔文在研究植物向性运动时,发现植物胚芽鞘的尖端受单侧光刺激后,向下面的伸长区传递了某种“影响”,造成伸长区背光面比向光面生长快,胚芽鞘出现向光性弯曲(图1)。这就是中学生物教科书上被大家广为熟知的达尔文向光性实验。1928年,荷兰科学家温特证实胚芽鞘弯曲生长是由一类化学物质引起的,并命名为生长素(auxin)。1946年,科学家从高等植物中首次分离出生长素,其主要成分为吲哚乙酸IAA。
后续研究发现,生长素不仅与植物向光性相关,还与植物向地性(向重力性)、向化性(包括向肥性)等相关。植物受单向的环境刺激而呈现的定向反应统称为向性(tropism)。这种向性主要是由生长素在植物体内极性分配造成的。因此,生长素的极性运输在这一过程中就变得非常关键。
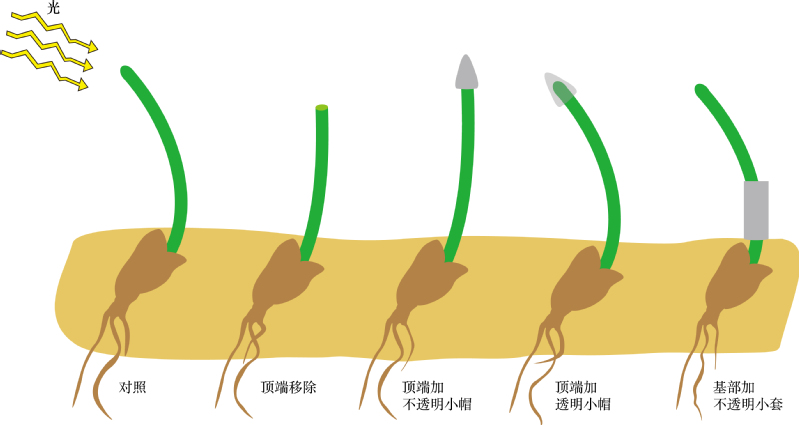
达尔文植物向光性实验示意图
生长素极性运输主要依赖于三种定位于细胞膜上的转运蛋白:AUX/LAX家族蛋白、PIN家族蛋白和ABCB家族蛋白,其中生长素外排蛋白PIN起最主要作用。pin突变体通常表现出相应组织生长素极性运输缺陷的表型,如向光性、向重力性受损等。然而,由于缺乏结构信息,人们对PIN蛋白如何介导生长素外排了解甚少。
PIN如何识别和转运生长素?
浙大研究团队利用单颗粒冷冻电镜技术,解析了拟南芥PIN3(AtPIN3)在未结合配体(AtPIN3apo)和结合IAA(AtPIN3IAA)两种状态的高分辨率结构。通过结构解析发现AtPIN3以二聚体形式存在,每个亚基包含10个跨膜螺旋(TM1–10),TM1–5和TM6–10组成反向重复结构(图2a)。AtPIN3apo与AtPIN3IAA结构类似,且均为向内开放状态。AtPIN3单体由支架结构域(scaffold domain)和转运结构域(transport domain)组成。在AtPIN3IAA结构中,IAA结合在支架结构域和转运结构域之间的内向开放口袋中(图2b),与多个保守的氨基酸发生氢键和疏水相互作用(图2c)
为了确定IAA与AtPIN3的结合模式,研究团队建立了体外放射性3H-IAA转运体系。相比于野生型AtPIN3,结合位点突变的3H-IAA外排活性显著降低。同时通过表面等离子体共振实验,测定野生型AtPIN3与突变体对IAA的解离常数(KD)。实验表明,野生型AtPIN3与突变体相比对IAA有更高的亲和力。此外,本研究同时结合了分子动力学模拟和结构比较等方法对IAA的结合模式进行多重验证,阐明了AtPIN3对IAA的识别与转运机制。
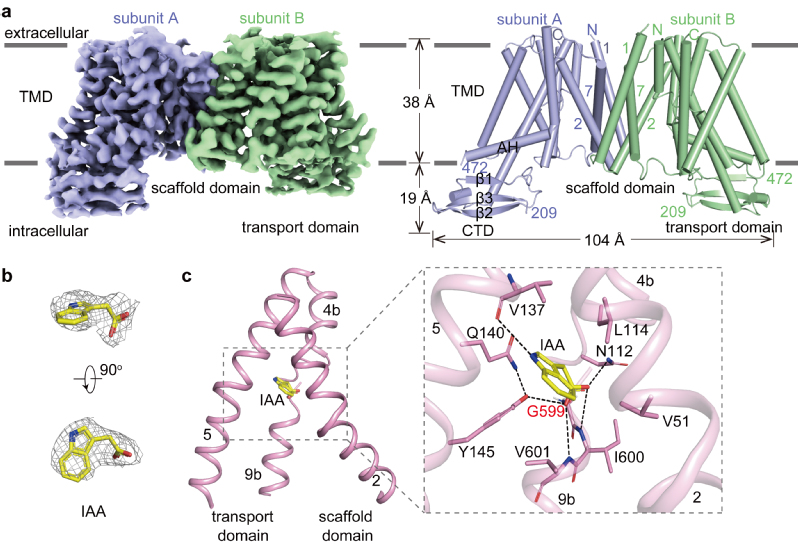
AtPIN3IAA结构。a. AtPIN3IAA的密度图和结构图。b. IAA的密度图。
c. AtPIN3IAA结构中IAA的结合位点。
NPA如何抑制生长素极性运输?
除草剂NPA(N-1-naphthylphthalamic acid)是一种被广泛研究的生长素极性运输抑制剂。NPA处理的植株几乎跟生长素转运蛋白突变体植株pin1的表型一模一样。长期以来,人们对NPA的作用靶点和作用机制一直存在争议。
该研究团队进一步解析了AtPIN3与抑制剂NPA复合物结构(AtPIN3NPA)(图3a,b)。研究人员同样通过体外放射性3H-IAA转运实验和表面等离子体共振实验验证了NPA的结合位点。比较AtPIN3IAA和AtPIN3NPA发现,抑制剂NPA与底物IAA的结合位点重叠(图3c)。因此,NPA为竞争性抑制剂,直接占据了PIN中生长素的结合位点,抑制了转运过程中PIN的潜在构象变化。
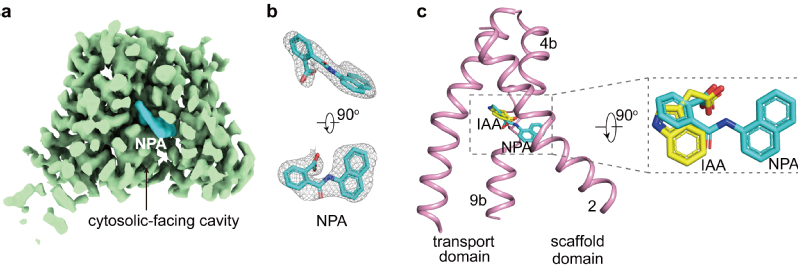
AtPIN3NPA结构。a. AtPIN3NPA的密度图。
b. NPA的密度图。c. IAA与NPA结合位点叠合。
基于上述结构和活性分析,研究团队提出了PIN介导生长素转运和NPA抑制生长素极性运输的分子机制。PIN处于内向开放状态时,细胞内的IAA结合在内向开放口袋中,引起PIN二体由内向开放状态向外向开放状态转换,IAA被释放至细胞外(图4)。抑制剂NPA结合在底物结合位点,阻碍了IAA的结合,同时抑制转运过程中PIN的潜在构象变化,起到抑制生长素极性运输的效果(图4)。
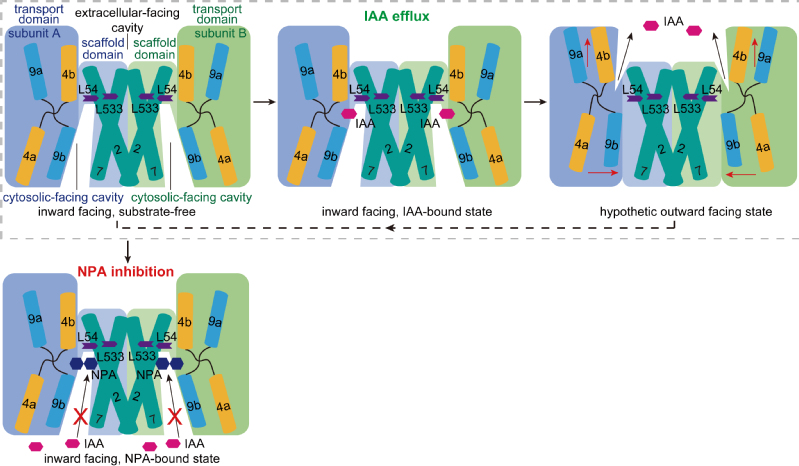
AtPIN3转运IAA和NPA抑制AtPIN3的模型
该研究解析了3个AtPIN3高分辨率电镜结构,明确了底物IAA和抑制剂NPA与AtPIN3的结合模式。这项工作不仅阐明了人们长久以来期待的PIN介导生长素转运的分子机制,而且将有助于进行作物改良,指导新型PIN抑制剂的开发。这些抑制剂既可作为生长素极性运输机理研究的工具,也可作为农业除草剂,具有广泛的应用前景。
该工作是由浙江大学、湖北大学和天津大学的多个实验室联合完成(图5)。浙江大学医学院郭江涛组博士后苏楠楠、杨帆组博士生竺爱琴和湖北大学生命科学学院吴姗组博士生陶鑫为论文的共同第一作者,郭江涛、吴姗和杨帆为共同通讯作者。参与这项工作的还有浙江大学生命科学学院郑绍建教授和丁忠杰研究员、浙江大学冷冻电镜中心常圣海老师、浙江大学农业与生物技术学院郭逸蓉老师和张素芬老师、天津大学生命科学学院叶升教授和湖北大学生命科学学院马立新教授。

浙江大学研究团队主要成员合影。前排左起:郭逸蓉、张素芬、张艳、苏楠楠、竺爱琴、杨帆;
后排左起:周晨羽、叶繁、郑绍建、郭江涛、常圣海。
(图文由课题组提供)

