
生物利用包括葡萄糖在内的各种糖类和脂肪作为碳源和能量的来源。根据被利用的顺序,碳源可以分为优先利用碳源(如葡萄糖)和非优先利用碳源。碳分解代谢物阻遏(carbon catabolite repression,CCR)是调控碳源利用顺序的一种机制。稻瘟菌(Magnaporthe oryzae)是一种引起水稻严重病害的真菌。在侵染前,稻瘟菌利用自身储存的脂滴、海藻糖和糖原等碳源;在侵染后,稻瘟菌经过活体营养和死体营养2个阶段,分别利用不同类型的植物碳源。课题组前期报道:AMP激活的蛋白激酶(AMPK)Snf1复合体是稻瘟菌利用脂肪和乙酸盐必需的(Zeng et al., 2014);C2H2转录因子CreA是脂肪分解代谢的负调控因子(Cao et al., 2016);bHLH转录因子Crf1是脂肪、甘油、阿拉伯糖和乙醇等多种碳源利用必需的,而且Crf1和CreA协同调控脂肪代谢(Cao et al., 2018);丝氨酸/苏氨酸蛋白磷酸酶Pp4调节亚基Smek1调控稻瘟菌的脂肪代谢和糖代谢(Huang et al., 2023)。Smek1通过去磷酸化CreA或者去磷酸化Crf1协调稻瘟菌对葡萄糖、橄榄油和阿拉伯糖(植物细胞壁的一种重要碳源)等各种碳源的利用。Smek1也是第一个在真核生物中被证实的调控甘油三酯分解代谢的蛋白磷酸酶。尽管如此,目前还没有建立关于真菌的CCR和CCDR(carbon catabolite de-repression)的比较完整的调控网络。

2025年1月28日,浙江大学生命科学学院卢建平课题组在Communications biology杂志上在线发表了题为 “A multilayered regulatory network mediated by protein phosphatase 4 controls carbon catabolite repression and de-repression in Magnaporthe oryzae”的研究论文。该研究揭示了稻瘟菌在利用宿主植物不同类型碳源时的代谢基因表达的调控机制,阐明了丝状真菌CCR和CCDR的多层次的调控网络。
首先,本研究确认了蛋白磷酸酶Pp4催化亚基Pp4c是与调节亚基Smek1互作的5个蛋白磷酸酶催化亚基之一,而且Pp4c和AMPK蛋白激酶Snf1都与转录抑制因子CreA和转录促进因子Crf1存在直接的体内互作关系。Pp4c是稻瘟菌生长、产孢、孢子萌发、附着胞穿透植物细胞壁和致病必需的。敲除PP4C导致该菌对葡萄糖、木糖、阿拉伯糖、橄榄油、甘油和乙醇的利用能力下降。通过比较单敲除和双敲除基因突变体之间的碳源利用能力,确认:Pp4c、Smek1、Snf1和CreA是病菌利用葡萄糖必需的,而Pp4c、Smek1、Snf1和Crf1是利用甘油三酯和阿拉伯糖必需的。通过比较在葡萄糖、橄榄油和阿拉伯糖培养基中相关代谢基因的表达水平,验证了这5个调控因子在甘油三酯分解代谢(脂解、β-氧化和糖原异生)和阿拉伯糖代谢中的作用位点和功能(图1)。
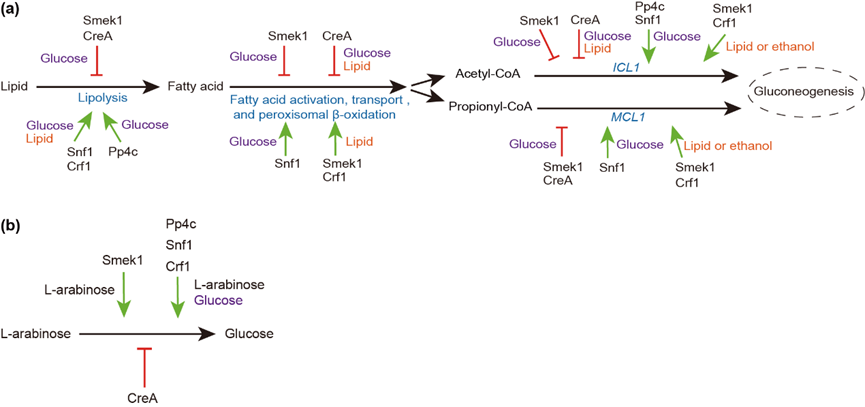
图1:Pp4c、Smek1、Snf1、CreA和Crf1在稻瘟菌脂肪分解代谢(a)和阿拉伯糖代谢(b)中的调控作用
进而,通过蛋白互作和蛋白磷酸化分析,确认:病菌利用葡萄糖时,Snf1磷酸化CreA和Crf1。利用阿拉伯糖时,Snf1磷酸化Pp4c,而磷酸化的Pp4c去磷酸化CreA和Crf1;同时,Smek1和磷酸化的Pp4c协同去磷酸化Snf1。结合课题组前期的研究结果,提出了一个关于稻瘟菌在利用葡萄糖(优先利用碳源)和阿拉伯糖(非优先利用碳源)时的代谢调控网络(图2)。
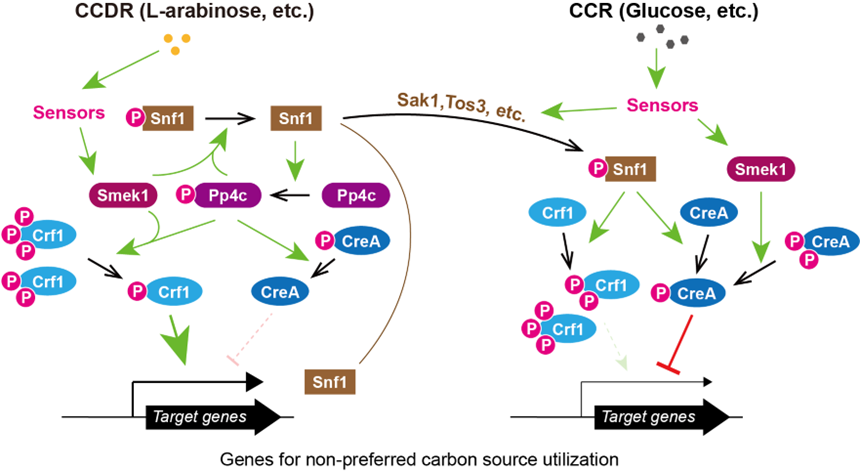
图2:稻瘟菌的碳分解代谢物阻遏与解阻遏调控网络
总之,该研究构建了一个调控稻瘟菌碳分解代谢物阻遏与解阻遏的网络骨架。当葡萄糖被用作碳源时(CCR条件),磷酸化的蛋白激酶Snf1和蛋白磷酸酶Smek1共同调节转录因子CreA和Crf1的磷酸化状态,导致CreA激活和Crf1失活,抑制参与非优先碳源分解代谢的基因的表达以及非优先碳源的利用。在CCDR条件下(例如,当使用阿拉伯糖作为碳源时),Snf1使蛋白磷酸酶Pp4c发生磷酸化,磷酸化的Pp4c和 Smek1随后使CreA和Crf1去磷酸化,导致CreA失活和Crf1激活,促进参与非优先碳源分解代谢的基因的表达以及非优先碳源的利用(图2)。鉴于Pp4和AMPK在包括植物和动物在内的真核生物中非常保守,该研究结果也为深入理解动植物的脂肪代谢、糖代谢以及肥胖症的调控途径提供了线索。
本文的第一作者为浙江大学生命科学学院毕业博士研究生黄志成,卢建平教授为通讯作者。浙江省农业科学院林福呈教授、王教瑜研究员和浙江大学农业与生物技术学院刘小红教授等参与了该研究。

